

How 3.5 billion years of cellular engineering maps to AI agent architecture — and why the math works
Multi-agent AI systems fail in predictable ways: runaway loops, hallucination cascades, prompt injection, token exhaustion. These aren't novel engineering problems. They're the same coordination failures that biological cells solved billions of years ago through network topology — specific wiring patterns that guarantee stability regardless of what the individual components are doing.
I've been building a framework called Operon that applies these biological motifs to AI agent systems. This post is an overview of the architecture: the mapping between biology and software, the key mechanisms, and the category theory that makes it more than a metaphor.
The Core Insight: Genes and Agents Are the Same Object
In a biological cell, thousands of genes act as autonomous agents. Each gene reads local chemical signals (its context), expresses proteins (its actions), and those proteins regulate other genes. The system is distributed, stochastic, noisy, and — mostly — it works. Not because the individual genes are perfect, but because the wiring between them enforces stability.
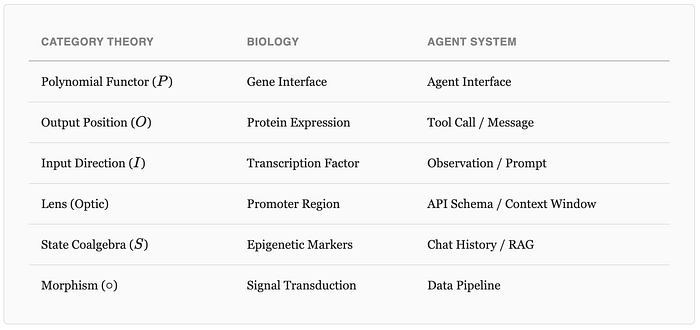
The mathematical object that captures both genes and agents is the polynomial functor, from applied category theory:

In plain English: a system that can be in various output positions (O), and for each output, accepts a specific set of inputs (I). A gene outputs proteins and accepts transcription factors. An agent outputs messages/actions and accepts observations/prompts. Same structure, different substrate.
This isn't just an analogy. When you formalize both systems as polynomial functors, you can prove things about agent architectures using results from gene regulatory network theory. The topology — how agents are wired together — determines the system's stability properties, independent of what the individual agents do.
The Six Organelles
A biological cell isn't just genes. It has shared infrastructure — organelles — that every gene relies on. Operon implements six software organelles, each mapped to a biological counterpart.
Membrane: Input Filtering (Immune System)
The cell membrane distinguishes Self from Non-Self. It runs two defense systems: innate immunity (fast, pattern-based — Toll-like receptors detecting conserved pathogen signatures) and adaptive immunity (slow, learned — T-cells trained to detect novel threats through behavioral profiling).
In Operon, the Membrane filters agent inputs against prompt injection, jailbreaks, and adversarial payloads. The innate layer matches known attack patterns (regex-based TLR patterns). The adaptive layer profiles agent behavior over time and flags statistical anomalies — if an agent's response patterns drift from its trained baseline, it may be compromised.
The math: we define a Provenance Functor that assigns trust levels to messages based on their structural origin (User, Tool, Agent_Self, Retrieved), not their content. A Trust-Gated Lens is a partial lens where the put operation only succeeds if the message's provenance meets the required trust threshold. Content-based attacks can't elevate provenance because the functor operates on message flow structure, not content.
Mitochondria: Safe Computation (MIPS)
Mitochondria aren't just the cell's powerhouse — recent research frames them as a Mitochondrial Information Processing System. They sense environmental stress, integrate metabolic signals, and govern cellular decisions.
In Operon, the Mitochondria is an AST-based safe computation engine. Arithmetic, trigonometry, boolean logic, JSON transforms — all on CPU, all sandboxed, all with capability-based access control. No tokens burned for deterministic computation. No injection risk from dynamic code execution.
Chaperone: Output Validation (Protein Folding)
Chaperone proteins (GroEL/GroES) cage misfolded proteins and give them multiple chances to fold correctly. If they can't fold, they're tagged for degradation.
In Operon, the Chaperone forces raw LLM output into strictly typed Pydantic schemas through a multi-strategy cascade: STRICT, EXTRACTION, LENIENT, REPAIR. Each strategy produces a confidence score. If all strategies fail, the output gets ubiquitin-tagged (marked for degradation). The Chaperone Loop extends this into a feedback loop: validation errors are passed back to the generator as context, enabling targeted repair rather than blind retry.
The math: the Chaperone is a partial retraction. The healing loop is a coalgebra with state S = Output x ErrorTrace and structure map that either succeeds or retries with error context.
Nucleus: LLM Integration Hub
The cell nucleus houses DNA and controls transcription. Operon's Nucleus wraps LLM providers (Anthropic, OpenAI, Gemini) with auto-detection, fallback, tool integration, and audit trails. The nuclear envelope abstracts provider complexity behind a consistent interface.
Ribosome: Prompt Template Engine
Ribosomes read mRNA and synthesize proteins via tRNA. Operon's Ribosome reads prompt templates and synthesizes concrete prompts via variable bindings — the same read-template-produce-output structure.
Lysosome: Cleanup and Recycling
Lysosomes digest cellular waste through autophagy. Operon's Lysosome handles failed operations, expired cache, and sensitive data disposal. It extracts debugging information from failures (recycling) and securely destroys PII and credentials (toxic disposal).
Network Motifs: Safety from Topology
The organelles handle individual agent health. Network motifs handle how agents interact — and this is where the real safety guarantees come from. Biology discovered that specific wiring patterns suppress errors regardless of what flows through the wires.
Coherent Feed-Forward Loop (CFFL): Two-Key Execution
An action proceeds only if both an executor and an independent risk assessor approve — an AND gate in the wiring.
The math: if the generator's error probability is p and the verifier's is q, and errors are independent, then

But LLM errors are often correlated (same training data, similar blind spots). With correlation coefficient ρ:

Same model, same prompt: ρ near 1, minimal benefit. Different model families: ρ in [0, 0.3], strong benefit. LLM + symbolic verifier: ρ near 0, maximum benefit. The topology provides multiplicative error suppression, but component diversity determines the actual coefficient.
Quorum Sensing: Multi-Agent Consensus
Bacteria coordinate through diffusible signaling molecules. When population density exceeds a threshold, they act collectively (bioluminescence, biofilm formation). Operon's Quorum Sensing aggregates votes from N agents. The final action fires only when signal concentration exceeds a confidence threshold. Seven voting strategies from simple majority to weighted consensus.
Morphogen Gradients: Coordination Without Central Control
In embryonic development, morphogens are diffusible signals. Cells read local concentration and differentiate accordingly. No blueprint, no central controller — just local sensing.
In Operon, morphogen gradients are shared context variables (COMPLEXITY, CONFIDENCE, BUDGET, ERROR_RATE, URGENCY, RISK) that agents read and respond to. High COMPLEXITY leads to detailed reasoning. Low BUDGET triggers graceful capability reduction. Low CONFIDENCE triggers quorum sensing. Agents adapt their phenotype to local conditions without being told what to do.
Oscillators: Periodic Rhythms
Circadian clocks, heartbeats, cell cycles — biology generates internal rhythms through delayed negative feedback. Operon's oscillators handle periodic health checks, context pruning, and maintenance tasks. Most agent systems are purely reactive; biological systems are proactively self-maintaining.
State Management: Epigenetics for Agents
Genes don't just react to the present — they carry memory through epigenetic markers that don't change the DNA but change how it's expressed. Operon implements three state management systems inspired by this.
HistoneStore provides multi-type memory with decay and reinforcement. Methylation markers are permanent (learned preferences). Acetylation markers are temporary (session context). Phosphorylation markers are dynamic (real-time state). Temporary memories fade over time; important ones get reinforced. Memories can be inherited by child agents.
ATP_Store manages multi-currency energy budgets. ATP for general operations, GTP for specialized tool calls, NADH as convertible reserves. This enforces termination: if every non-identity transition costs at least c_min > 0 and the total budget is R, then every execution terminates in at most floor(R / c_min) steps. Termination becomes decidable.
Telomere tracks agent lifecycle. Telomeres shorten with each cell division; critically short telomeres trigger senescence or apoptosis. Operon's Telomere counts operations: BIRTH, GROWTH, MATURITY, SENESCENCE, APOPTOSIS. Agents age, degrade gracefully, and die cleanly.
Self-Repair: Three Healing Modalities
Biological cells don't just detect failures — they continuously repair themselves. Operon implements three healing mechanisms corresponding to structural, metabolic, and cognitive damage.
Chaperone Loop (Structural Healing): Feed validation errors back to the generator as context. The error trace itself is information — "TypeError: 'one hundred' is not a valid float" tells the generator exactly what to fix. Confidence decays with each retry. If all attempts fail, ubiquitin-tag for degradation.
Regenerative Swarm (Metabolic Healing): Detect stuck workers via entropy monitoring (repeated outputs = no progress). Kill the stuck worker (apoptosis), summarize what it tried, respawn a replacement with the hint: "Worker_1 failed trying strategy X. Try a different approach." The dying cell's debris signals its successors.
Autophagy Daemon (Cognitive Healing): Monitor context window utilization. When it exceeds a threshold, the agent enters a sleep cycle: useful state is summarized into long-term memory (histone markers), raw context is flushed, and the agent resumes with a clean window plus summary. This prevents the context equivalent of protein aggregate buildup.
Why Category Theory?
You could build all of this without the math. The biological metaphors alone generate useful engineering patterns. So why bother with polynomial functors and operads?
Three reasons.
Transferable results. If two systems are modeled by the same polynomial functor, results proven for one transfer to the other. Gene regulatory network stability theorems become agent coordination stability theorems. This is what makes the biology more than a metaphor — it's a source of proven architectural patterns.
Composition guarantees. The Agentic Operad (WAgent) defines which agent wirings are valid at the type level. If Agent A outputs natural language but Agent B expects JSON, the composition is undefined — a compile-time error, not a runtime crash. You can reason about the properties of a composite system from the properties of its components. Biology's typed wiring (specific transcription factors bind specific promoters) maps directly to this.
Provable bounds. The Metabolic Coalgebra gives you a termination guarantee: with finite budget and positive minimum cost per transition, every execution terminates. This is a theorem, not a hope. The CFFL error suppression bound is quantitative — you can compute the actual safety gain for a given correlation coefficient between your agents.
The framework draws on Spivak's work on polynomial functors, Vagner's algebras of open dynamical systems, and Friston's Free Energy Principle (for the Epiplexity monitor). The full formalization is in the accompanying paper.
The Bet
Operon's bet is simple: if you apply the same topological patterns that keep cells alive to AI agent systems, those systems inherit cellular robustness. Safety emerges from structure, not from prompting. A cell doesn't stay healthy because each gene individually decides to behave — it stays healthy because the wiring between genes enforces homeostasis. Agent systems can work the same way.
I've been building these biological motifs into a Python library with 22 interactive demos you can try in-browser — no setup, no API keys. If you want to see the cancer detector (Epiplexity Monitor), the immune system (Membrane + Innate Immunity), the ATP budget, or the chaperone healing loop in action: huggingface.co/coredipper. Code at github.com/coredipper/operon.